
|
IN THE BOHEMIAN SILURIAN AND LOWER DEVONIAN. (2 text-figs, Pl. I-VII)
|
Rudolf J. Prokop - Václav Petr
Key words: Crinoidea, Flexibilia, Lecanocrinus, Silurian, Lower Devonian, Barrandian, Prague Basin, Czech Republic, Systematics, Biostratigraphy, Phylogeny.
Abstract: Skeletal remains of the crinoid genus
Lecanocrinus HALL, 1852 are described from the Silurian (Ludlow
- Ludfordian) and Devonian (Lochkovian, Pragian and Zlíchovian) limestone
facies of the Barrandian area for the first time. They belong to the following
species: Lecanocrinus facietatus (ANGELIN, 1878) - the only known
Silurian species from Bohemia, L. cf. facietatus (ANGELIN,
1878) - from the Lochkovian, L. sp. and L. soyei OEHLERT,
1882 - from the Pragian, and L. hanusi sp. n. - from the Zlíchovian.
The crinoid Lecanocrinus HALL, 1852 represent possibly an exception among the relatively rare members of the subclass Flexibilia. Its numerous species have been found in rocks of the Silurian (Wenlock) to the Devonian (Eifelian) age in England, Gotland (Sweden), Germany, France, United States and Australia. Especially in the Ludlowian rocks it is known to be a cosmopolitan genus (WITZKE, FREST & STRIMPLE, 1979). In this article various isolated skeletal elements (rarely more or less complete calyces and incomplete arms) of Lecanocrinus are reported also from the Barrandian area. The most complete remains have been discovered especially in the Ludlow (Upper Silurian, Kopanina Formation, trilobite horizon with Ananaspis fecunda) and in the Zlíchovian (Lower Devonian, base of the Zl?chov Formation, the so-called "Chapel Coral Horizon"). In the Lower Devonian of the Barrandian area the isolated skeletal elements (predominantly BB, RR, and BrBr) of this genus are relatively frequent but the presen ce of Lecanocrinus in Bohemia was only briefly reported in articles of several previous authors (Hanuš 1923, 1927; Bouška 1946, 1956; Prokop 1987, 1992). No descriptions or figures have been given.
All the Bohemian species would belong to Lecanocrinus s. s., i.e. to the typical subgenus Lecanocrinus (L.) as defined by Frest et Strimple, 1978. It is important to point out that in many respects (not in all!) the general phylogenetic trend among the numerous species of L. (L.) leading to the related younger genus Geroldicrinus Jaekel, 1918 (Middle Devonian of Germany) has been observed during our study of the material from the Barrandian area. However, because of some exceptions from this trend (mentioned in the systematic part) it seems to us to be useful to prefer herein a simple classification of the genus Lecanocrinus, i.e. without the establishment of the subgenera. The phylogenetic trend in all lineages of the subfamily Lecanocrininae (including acquisition of basal concavity and reduction in arm length and branching) as outlined in their thorough revision by Frest et Strimple (1978) must be taken only as a broad generalization. It is long known that there is no orthogenesis or anagen esis in evolution (see e.g. Schindewolf, 1957).
Almost all material has been obtained by washing of the weathered parts of Barrandian limestones (i.e. from the so-called "white beds"). Unquestionable remains of this genus have been found in limestone layers of all Lower Devonian stages of the Barrandian area except the Dalejan (corresponding to Upper Emsian). In the Dalejan strata only small, thin isolated RR have been ascertained (RP) but their relation to the genus Lecanocrinus Hall, 1852 is doubtful. Therefore, the latter are only figured but not described in this article. All material is deposited in collections of the Palaeontological Department of the National Museum (Museum of Natural History), Praha.
We are indebted to Dr. V. Turek (National Museum) for making all the
photographs in this article. We gratefully acknowledge valuable comments
of Prof. I. Chlupáč (Charles University) who also deserves special thanks
for the beautiful specimen of Lecanocrinus coming from the eastern wall
of "Císařský lom" quarry near Koněprusy.
Lecanocrinus HALL, 1852
Type species: Lecanocrinus macropetalus HALL, 1852; Upper Silurian, New York, U.S.A.
Stratigraphic and palaeogeographic distribution: Silurian (Wenlock) to Devonian (Eifelian) of Gotland (Sweden), England, Germany, France, Australia, U.S.A. Although in the world the great majority of species comes from Silurian rocks, the reverse situation is obviously encountered in Bohemia where only one less common species (although almost complete calyx) was discovered in the Silurian strata.
Lecanocrinus facietatus (ANGELIN, 1878)
(Pl.I, fig. 1-3)
1878 Cyrtidocrinus facietatus Angelin; Angelin, p. 20, Pl. 21,
fig. 13, 14, 14a.
1880 Cyrtidocrinus facietatus Angelin; Wachsmuth et Springer,
pt. I, p. 145.
1920 Lecanocrinus facietatus (Angelin); Springer, p.133-134,
Pl. II, fig. 16-21.
Material: 1 complete calyx (L 30565), figured herein on Pl.I, figs. 1-2, and 2 isolated RR.
Description of the calyx from "Mušlovka": Calyx relatively broadly conical, widest at the summit of RR, with thin plates. The outer surface of the plates is convex, smooth, with remains of a tiny granulation. IBB circlet very low but visible from the side view. BB large, the ratio of the height of BB to that of RR is about 5:6. The RA is slightly smaller than the anal X and like the DR it is poorly preserved. The well-preserved anal X is relatively large, hexagonal, widest proximally, and it does n ot reach above the summit of the RR while the CR and DR do not touch each other. The pentagonal RR bear plenary radial facets with distinct transverse rim and relatively short aboral ligament fossa. Arms not preserved. There is a barely concave basal pit with the subcircular to subpentagonal proximal columnal present, bearing a tiny central lumen and short but distinct crenellae on the margin.
Remarks: No important differences from the specimens described by Springer (1920) from Gotland have been found. For other remarks see those on Lecanocrinus hanusi sp. n.
Dimensions of the calyx from "Mušlovka" (in mm.):
height of the calyx 9.0
width of the calyx at the summit of RR 11.5
height of the IBB circlet 1.0
width of the IBB circlet 5.5
width of the BB circlet 10.0
height of the anal X 4.5
diameter of the stem facet 3.2
Occurrence: All Bohemian material was derived by B. Bouček from
the washings in the trilobite horizon with Ananaspis fecunda at
the quarry "Mušlovka" at Praha 5 - Řeporyje, Kopanina Formation (Ludlow,
Ludfordian - stratigraphic terminology after Kříž, 1992).
Lecanocrinus cf. facietatus (ANGELIN, 1878)
(Pl. I., figs. 4-11)
Material: One incomplete calyx with dislocated plates (L 29079) preserved on the weathered part of a limestone sample (see Pl.I, fig. 4) and several isolated RR and PBrBr.
Description: The preserved anterior part of the calyx is laterally compressed and the plates are partly dislocated. Calyx was probably bowl-shaped, with slightly vaulting cup plates. There are only 3 BB and 4 RR present, all of them are distinctly thin-walled and smooth. Although the IBB circlet is not preserved, the rounded shape of lower (proximal) edges of the subpentagonal BB suggests that there were IBB present and that they were at least slightly visible from the side view. The BB are about two times smaller than the pentagonal RR. The radial facets are plenary, with exceptionally marked transverse rim and with relatively slender but deep aboral ligament fossa. In the specimen L 29079 also two low, dislocated PBrBr are preserved and proximal part of the stem - again, with very slightly disarticulated columnals. These proximal columnals are very thin, circular, with tiny rounded lumen and broad areola. The crenelation present on the margin of the columnals is short but extremely prominent , pointing to the symplectial type of columnal articulation at least in the proxistele.
Occurrence in the Kotýs Limestone (Lower Devonian, Lochkovian, Lochkov Formation):
a) Svatý Jan pod Skalou, quarry "Na Stydlých vodách" (calyx L 29079);
b) Praha-Řeporyje, quarry "Černá skála" (isolated RR);
c) quarry "Na Čížovci" near Trněný Újezd (isolated RR);
d) Loděnice near Beroun, quarries "Na Branžovech" ("Záloženský" quarry)
(isolated RR and BrBr).
1882 Lecanocrinus soyei n. sp.; Oehlert, p. 354, Pl. VIII, fig.
2.
1920 Lecanocrinus soyei Oehlert; Springer, p. 142-143, Pl. III,
figs. 27-29.
1985 Lecanocrinus soyei Oehlert; Le Menn, p. 104-107, text-fig.
43, Pl. 21, figs. 13-15; Pl. 22, figs. 1-7.
Material: one complete base of the calyx with RA attached (L 29080, Pl. II, figs. 1-5) and many tens of isolated skeletal elements (especially BB, RR and BrBr) derived from washings of the "white beds" of limestone facies listed below.
Description: Proximal part of the calyx is cup-shaped, with barely concave basal pit. IBB circlet is low but visible from the side view. BB relatively high, elongated, pentagonal. RA is rectangular, almost square-shaped, relatively large, forming about 1/2 of the size of a basal. RR are known only as isolated plates, they are thick-walled, pentagonal, their ratio height:width fluctuates from 1:1 (in small specimens) to 1:1.4 (in normal adults). Identifiable anal X at our disposal (although only isolated). CR and DR are also clearly present in the material, being laterally concave and drawn along their upper (distal) margin into moderately sharp edges, suggesting connection with anal X which very probably did not reach above the summit of RR (as in the specimens from France). Radial facets are plenary, bearing distinct transverse rim and a broad, deep aboral ligament fossa. In well-preserved specimens the fossa is crenelated. The outer surface of RR is covered with dense, distinct granulation w hich in some places (especially in the proximal part of the plates) is marked with gradation into vermiculate structures.
Remarks: The above described skeletal elements are practically identical with those discovered in the Gedinnian and Siegenian (corresp. to the Lochkovian and Pragian) of France (see especially Le Menn, 1985, Pl. 21, 22).
Dimensions of the proximal part (IBB, BB and RA) of the aboral cup from Dvorce-Prokop Limestone (in mm.):
height 7.0
maximal width 12.5
height of IBB 0.2
diameter of the basal concavity 4.5
Occurrence in the Bohemian Praha Formation (Lower Devonian, Pragian):
1. Slivenec Limestone:
a) Praha-Smíchov, quarry "Na Konvářce",
b) Srbsko (Beroun district), quarry in the Kačák Valley near the place
where the brook empties itself into the Berounka River;
2. Loděnice Limestone:
a) Praha-Smíchov, quarry "Na Konvářce",
b) Praha-Klukovice, western part of the "Červený lom" quarry.
3. Dvorce-Prokop Limestone:
a) Praha-Malá Chuchle, quarry near the wood road to Slivenec,
b) Praha-Hlubočepy, quarry near St. Prokop,
c) Praha-Klukovice, eastern part of the "Červený lom" quarry, and
d) Praha-Řeporyje, quarry "U kantiny" (here the washings come from
limestone layers 90 to 2 cm below the base of the Zlíchov Limestone);
Lecanocrinus sp.
(Pl. V., fig. 1)
Material: One almost complete crown with partly incomplete base and arms (L 12106, figured herein on Pl. V., fig. 1) preserved on weathered surface of bioclastic limestone, and some isolated RR.
Description: Poorly known species. The only known calyx is broadly conical, with straight walls, its height is 5.5 mm, while the width reaches 9.0 mm. It is composed of thick, weathered thecal plates which are smooth; in unweathered portions of arms there is however markedly visible granulation. IBB partly preserved, low, but on one side they are visible from the lateral view. BB also low and broad. The preserved quadrangular RA forms about 1/2 of the size of a radial plate. It is interesting that the RA is so low situated that its lower margin is in contact with the uppermost (distal) edge of the IBB. The anal X is of the shape of high hexagon, attaining almost one half of the width of a radial plate. Important feature of the anal X is that it does not reach above the summit of RR. The large, pentagonal RR have slightly concave plenary radial facets with arms attached to them in their whole width. The observable part of the arms, which are turned inward, branches three times. BrBr are relat ively high, number of PBrBr is 1-2, SBrBr 2-3, number of TBrBr (3 visible) is uncertain. Stem is unknown. For remarks see those on the following species Lecanocrinus hanusi sp. n.
Occurrence: Lower Devonian, Pragian, Praha Formation, Koněprusy Limestone, bioclastic facies (talus zone of the Pragian reef) at the eastern wall (Zlatý kůň Hill) of the "Císařský lom" quarry near Koněprusy (calyx L 12106 found in 1969 by I. Chlupáč) and in the same facies in the quarry "Na Plešivci" near Suchomasty (only isolated RR).
Lecanocrinus hanusi sp. n.
(Pl. V, figs. 2-5; Pl. VI, figs. 1-10; Pl. VII, figs. 1-10)
(Text-figs. 1 a,b; 2)
Holotype: incomplete calyx (L 29075) with the most proximal part
of the stem attached, isolated from the weathered bioclastic limestone,
figured herein on Pl. V, figs. 2-5.
Type horizon: Lower Devonian, Zlíchovian (corresponding to Lower
Emsian), basal part of the Zlíchov Formation, the so-called "Chapel Coral
Horizon".
Type locality: Praha - Zlíchov, quarry "U kapličky"
Material: except the holotype one almost complete isolated arm
with the radial plate attached (paratype A, L 29076, figured herein on
Pl. VI, figs. 1-3, and on the schematic text-fig. 1b, which served also
as an important model for the reconstruction on the text-fig. 2); isolated
fragment of the middle portion of the arm with TBrBr and QBrBr (L 30564,
figured herein on Pl. VI, fig. 5); three other fragments of arms, and several
tens of isolated IBB, BB, RR and BrBr.
Text-Fig. 1: Lecanocrinus hanusi sp. n.
a) plate diagram of the calyx based on the holotype (L
29075). RR black.
b) schematic sketch of a radial plate with the proximal
part of arm attached, based on the paratype A (L 29076). R black, PBr Ax
and non-axillary BrBr white, SBr AxAx indicated by oblique cross-ruled
pattern, TBr AxAx dotted, QBr Ax obliquely lined.
a pathological stereom (not interTBr)
b pathological duplication of SBrBr.
Remarks: Lecanocrinus hanusi n. sp. is similar to L.
lindstroemi Springer, 1920 from the Silurian of Gotland and to L.
soyei Oehlert, 1882 in having the anal X reaching only the level of
the summit of RR. This is very important and unique feature among the large
number of species of the genus Lecanocrinus because in all other members
of this genus the anal X reaches more or less distinctly but clearly above
the upper edge of RR. Therefore, in this characteristic, these three species
and Lecanocrinus sp. described herein from the Koněprusy Limestone
appear to be closer to the related genus Geroldicrinus Jaekel, 1918
in which the anal X does not reach above the summit of RR in any case.
However, we are unable to explain whether such feature was of phylogenetic
importance or whether it was derived simply from similar ecological specialization
of the different lecanocrinid species: any conclusion seems to be highly
speculative. In our opinion, from the phylogenetic point of view, the main
differen ce between the genera Lecanocrinus and Geroldicrinus
is really in the different character of arms and the IBB circlet and that
in this respect the short arms (branching only twice isotomously) and the
restriction of the IBB circlet to the basal concavity can serve for the
genus Geroldicrinus as features of utmost taxonomic importance.
On the other hand, the possibility of very close relationship between Geroldicrinus
roemeri (Schultze, 1867) and Lecanocrinus hanusi n. sp. cannot
be excluded because of presence of relatively deep basal pit in the latter
(although the IBB are very good visible from the lateral view). Moreover,
because of their very low cups, both Lecanocrinus hanusi sp. n.
and Geroldicrinus can be regarded as morphologically "advanced",
similar in a generalized view much more to many Upper Carboniferous crinoid
genera than to Lower Palaeozoic ones (Moore et Laudon 1943, Frest et Strimple
1978).
The discussed new species differs from L. lindstroemi (Frest et Strimple,
1978 supposed the latter is the ancestor of Geroldicrinus also because
of having pentalobate cup) especially in relatively larger RA and in the
drop-like appearance of the anal X which (in the Bohemian species) is distally
pointed to a tip. Finally, L. hanusi is similar to L. lindstroemi,
L. soyei and to L. facietatus in having barely concave basal
pit. The last similarity, without both the knowledge of presence of the
latter two species in Bohemian rocks and of the existence of L. hanusi,
led Frest et Strimple (1978, p. 525) to the conclusion that L. facietatus
and L. soyei form a separate lineage. Now, from the same point of
view, it is quite possible to construct the phylogenetic lineage in another
way: L. facietatus - L. soyei - L. hanusi - G.
roemeri, while L. lindstroemi can be regarded as a separate
but parallel line. In fact, both explanations are logical but it is clear
that both cannot be correct. What is wrong?
At present, by an irony of fate we do not know much more: rather less,
because we know only that looking at the evolution from such very old theoretical
viewpoint (i.e. "orthogenetic" directionality in "progress", gradual anagenetic
transformation of whole species into another one or its cladogenetic transformation
into another two ones) like Frest and Strimple (1978, especially their
reconstruction of the phylogenetic tree of Lecanocrininae on page 531,
fig. 4), then every new finding of a new fossil Lecanocrinus must
change the lineages in their tree giving an illusion of its increased accuracy.
However, it does not seem to be the right solution of the problem. In fact,
we can never say with certainty that a particular lecanocrinid species
is the ancestor of another one. Even from the time of the peak of old neodarwinism
in the so-called "modern synthesis" it is known that "orthogenetic" trends
did not occur in evolution (see e.g. Simpson, 1953). Also other schools
proposed at that time new explanations (e.g. the mosaic evolution of Schindewolf,
1957). After the collapse of "modern synthesis", during the time of recent
victory of the concepts of punctuated equilibria (Eldredge et Gould, 1972)
and of the macroevolution, evolutionists generally hold that species have
overlapped in time and survived for long intervals unchanged (e.g. Stanley,
1982). After Gould et Eldredge (1977, p. 115) "Phyletic gradualism was
an a priori assertion from the start - it was never 'seen' in the rocks;
it expressed the cultural and political biases of 19th century liberalism."
Now the trends are generally regarded as changes in variance and the directional
progress simply as bias (see e.g. Benton, 1987; Gould, 1988). In other
words, at present almost nobody would agree with such a reconstruction
of phylogeny as is outlined in Frest et Strimple (1978); they are right
if we consider the trends as a broad generalization - but the individual
lineages are obviously unreal.
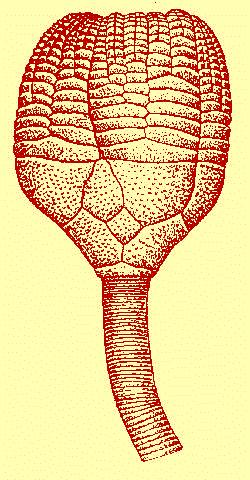
Text-Fig. 2: Lecanocrinus hanusi sp. n.
Reconstruction of the crown with proximal part of the
stem attached, based especially on the specimens L 29075, 29076 and 30564.
height of the calyx 9.0
width of the calyx at the summit of RR 16.0
height of the IBB circlet 1.5
width of the IBB circlet 6.5
width of the BB circlet 12.0
diameter of the stem facet 5.0
Occurrence: Relatively frequent in washings from the bioclastic limestones
of the so-called "Chapel Coral Horizon" at the base of the Zlíchov Formation.
Except the type locality it has been found in the layers of the same stratigraphical
position in the eastern part of the "?erven? lom" quarry near Praha - Klukovice.
ANGELIN, N. P. (1878): Iconographia crinoideorum in stratis Sueciae Siluricis fossilium. 62 pp., 29 pls. Samson & Wallin, Holmiae.
BENTON, M. J. (1987): Progress and Competition in Macroevolution. - Biol. Rev., 62: 305-338. Cambridge.
BOUŠKA, J. (1942): Krinoidengattung Pycnosaccus im böhmischen Silur. - Rozpr. Čes. Akad. Věd Umění, T?. II, 52(21): 1-4. Praha.
BOUŠKA, J. (1946): On Crotalocrinitidae (Angelin) from the Silurian and Devonian of Bohemia. - Rozpr. Čes. Akad. Věd Umění, T?. II, 56(4): 1-17. Praha.
BOUŠKA, J. (1956a): O rodu Protaxocrinus Springer (Crinoidea) ze st?edo?eského siluru. On the occurrence of the genus Protaxocrinus Springer (Crinoidea) in the Silurian of Bohemia. - Sbor. Ústř. Úst. geol., Odd. Paleont., 22(for 1955): 323-333. Praha.
BOUŠKA, J. (1956b): Pisocrinidae Angelin českého siluru a devonu (Crinoidea). - Rozpr. Ústř. Úst. geol., 20: 1-137 (1-54 Czech text, 55-96 Russian text, 97-137 English text). Praha.
BOWSHER, A. (1953): A new Devonian crinoid from western Maryland. - Smithsonian Misc. Coll., 121(9): 1-8. Washington, D. C.
BRETT, C. E. (1985): Tremichnus: a new ichnogenus of circular-parabolic pits in fossil echinoderms. - J. Paleont., 59(3): 625-635. Lawrence.
ELDREDGE, N. - GOULD, S. J. (1972): Punctuated equilibria: An alternative to phyletic gradualism. In: T. J. M. Schopf (Ed.): Models in paleobiology. 82-115, Freeman, San Francisco.
FREST, T. J. - STRIMPLE, H. L. (1978): A revision of Lecanocrinus (Crinoidea: Flexibilia) and a new North American Devonian lecanocrinid. - N. Jb. Geol. Paläont., Mh., 1978(9): 521-533. Stuttgart.
GOULD, S. J. (1988): Trends as changes in variance: A new slant on progress and directionality in evolution. - J. Paleont., 62(3): 319-329. Lawrence.
GOULD, S. J. - ELDREDGE, N. (1977): Punctuated equilibria: the tempo and mode of evolution reconsidered. - Paleobiology, 3(2): 115-151. Ithaca.
HANUŠ, F. (1923): O nových krinoidech (lilijicích) českého siluru a devonu. - Čas. Nár. Muz., Odd. přírodověd., 97: 38-39. Praha.
HANUŠ, F. (1927): Pravý poklad zůstatků devonské zvířeny u Prahy. - Čas. Nár. Muz., Odd. přírodověd., 101: 89-96. Praha.
JAEKEL, O. (1918): Phylogenie und System der Pelmatozoen. - Paläont. Z., 3(1): 1-128. Berlin.
KŘÍŽ, J. (1992): Prague Basin (Barrandian), Bohemia. In: M. G. Bassett (Ed.): Silurian Field Excursions. - National Museum of Wales, Geological Series Nr. 13, 111 pp. Cardiff.
LE MENN, J. (1985): Les Crinoides du Dévonien inférieur et moyen du Massif armoricain: Systématique - paléobiologie - evolution - paléontologie - biostratigraphie. - Mém. Soc. géol. minéral. Bretagne, 30: 1-268. Rennes.
McINTOSH, G. C. (1981): The crinoid Lecanocrinus Hall, 1852 (= Alsopocrinus Tansey, 1924) from the Lower Devonian of Missouri and Tennessee. - J. Paleont., 55(5): 962-966. Lawrence.
MOORE, R. C. (1978): Flexibilia. In: R. C. Moore & C. Teichert (Eds.): Treatise on Invertebrate Paleontology, Part T, Echinodermata 2, 2, T759-T812. The Geological Society of America, Boulder, Colorado, and the University of Kansas, Lawrence, Kansas.
MOORE, R. - LAUDON, L. (1943): Evolution and classification of Paleozoic crinoids. - Geol. Soc. Amer., Spec. Pap., 46: 1-153. Washington, D. C.
OEHLERT, D. P. (1882): Crinoides nouveaux du Dévonien de la Sarthe et de la Mayenne. - Bull. Soc. Géol. France, Sér.3, 10: 352-363, Pl. 8-9. Paris.
PROKOP, R. (1986): Contribution to crinoid stratigraphy. In: I. Chlupáč: Section below Praha - Barrandov, "U Kapličky" quarry (Pragian-Zlíchovian boundary), 15-18. In: Field Conference of the International Subcommission on Devonian Stratigraphy "Barrandian-Moravian Karst 1986": A field trip guidebook. - Ústř. Úst. geol., Praha.
PROKOP, R. (1986): Contribution to crinoid stratigraphy. In: I. Chlupáč: Červený lom quarry near Praha-Klukovice (Pragian-Zlíchovian boundary), 18-20. In: Field Conference of the International Subcommission on Devonian Stratigraphy "Barrandian-Moravian Karst 1986": A field trip guidebook. - Ústř. Úst. geol., Praha.
PROKOP, R. (1986): Contribution to crinoid stratigraphy. In: I. Chlupáč: Plešivec Hill near Měňany (Pragian fauna), 31-32. In: Field Conference of the International Subcommission on Devonian Stratigraphy "Barrandian-Moravian Karst 1986": A field trip guidebook. - Ústř. Úst. geol., Praha.
PROKOP, R. J. (1987): The stratigraphical distribution of Devonian crinoids in the Barrandian area (Czechoslovakia). - Newsl. Stratigr., 17(2): 101-107. Berlin, Stuttgart.
PROKOP, R. J. (1992): Newly ascertained genera of crinoids in the Devonian of Bohemia (Barrandian area). - Čas. Nár. Muz., Odd. přírodověd., 159 (for 1990) (1-4): 110. Praha.
PROKOP, R. J. - PETR, V. (1987): Eutaxocrinus capella sp.n. (Crinoidea, Flexibilia) from the Bohemian Lower Devonian (Zlichovian). - Čas. Mineral. Geol., 32(2): 165-167. Praha.
PROKOP, R. J. - PETR, V. (1989): New Silurian and Devonian crinoid taxa from Bohemia. - Čas. Mineral. Geol., 34(1): 45-57. Praha.
PROKOP, R. J. - PETR, V. (1991): Type specimens of fossils in the National Museum, Prague. Vol. 2: Palaeozoic Crinoidea.Palaeozoic Crinoidea. - National Museum, Museum of Natural History, Praha, pp.1-74, pl. I-X.
PROKOP, R. J. - TUREK, V. (1983): Unikátní nález silurských flexibilních krinoidů, přisedlých na schránku orthokonního nautiloida. A unique discovery of Silurian flexible crinoids attached to an orthocone nautiloid shell. - Čas. Nár. Muz., Ř. přírodověd., 152(4): 181-186. Praha.
SCHINDEWOLF, O. H. v. (1957): Über Mosaikentwicklung. - N. Jb. Geol. Paläont., Mh., 1957: 49-52. Stuttgart.
SCHULTZE, L. (1867): Monographie der Echinodermen des Eifler Kalkes. - Denkschr. Kais. Akad. Wiss., math.-naturwiss. Kl., 26(2): 113-230. Wien.
SIMPSON, G. G. (1953): The Major Features of Evolution. 434 pp., Columbia Univ. Press, New York.
SPRINGER, F. (1920): The Crinoidea Flexibilia. - Smithsonian Inst. Publ., 2501, 486 pp. Washington, D. C.
SPRINGER, F. (1926): American Silurian crinoids. - Smithsonian Inst., Publ. 2871, 239 pp., 33 pls. Washington, D. C.
STANLEY, S. M. (1982): Macroevolution and the fossil record. - Evolution, 36(3): 460-473. Lawrence.
STRIMPLE, H. L. (1952): New species of Lecanocrinus. - J. Washington Acad. Sci., 42(10): 318-323. Washington, D. C.
STRIMPLE, H. L. (1954): Two new crinoid species from the Henryhouse of Oklahoma. - J. Washington Acad. Sci., 44: 280-283. Washington, D. C.
STRIMPLE, H. L. (1963): Crinoids of the Hunton Group (Devonian-Silurian) of Oklahoma. - Oklahoma Geol. Surv. Bull., 100: 1-169. Norman.
WAAGEN, W. - JAHN, J. J. (1899): Classe des Echinodermes, 2, Famille des Crinoides. In: J. Barrande: Systeme silurien du centre de la Boheme, 7, 1-215. Praha.
WACHSMUTH, Ch. - SPRINGER, F. (1880): Revision of the Palaeocrinoidea, Pt. I: The families Ichthyocrinidae and Cyathocrinidae. - Proc. Acad. Nat. Sci. Philadelphia, 1879: 226-378, Pl. 15-17. Philadelphia.
WITZKE, B. J. - FREST, T. J. - STRIMPLE, H. L. (1979): Biogeography of the Silurian-Lower Devonian Echinoderms. In: J. Gray - A. J. Boucot (Eds.): Historical Biogeography, Plate Tectonics, and the Changing Environment, p. 117-129. Oregon St. Univ. Press, Eugene.
Explanations to the plates
Pl. I
1-3 Lecanocrinus facietatus (Angelin, 1878)
All specimens from the quarry "Mušlovka" at Praha - Řeporyje, Kopanina Formation (Ludlow, Ludfordian), trilobite horizon with Ananaspis fecunda.
1. posterolateral view of the holotype L 30565, x 3.2;
2. basal view of the same specimen, x 3.3;
3. isolated radial, L 30566, x 4.5;
4-11 Lecanocrinus cf. facietatus (Angelin, 1878)
All specimens from Kotýs Limestone (Lower Devonian, Lochkovian, Lochkov Formation); the cup on fig. 4 comes from Svatý Jan pod Skalou, quarry "Na Stydlých vodách", while 5-11 from the quarry "Černá skála" near Praha - Řeporyje.
4. lateral view of partly disarticulated cup L 29079,
x 3.1;
5. isolated R, L 30567, outer view, x 3.5;
6. isolated DR, L 30568, outer view, x 3.5;
7. isolated R, L 30569, inner view, x 3.5;
8. isolated R, L 30570, outer view, x 3.5;
9. isolated R, L 30571, inner view, x 3.5;
10. isolated PBr Ax, L 30572, distal view, x 3.5;
11. isolated PBr Ax, L 30573, distal view, x 3.5;
Pl. II
1-5 Lecanocrinus soyei Oehlert, 1882
The specimen L 29080 (only IBB, BB and RA preserved) from the Dvorce-Prokop Limestone (Lower Devonian, Pragian, Praha Formation) at Praha - Malá Chuchle, quarry near the wood road to Slivenec.
1. basal view, x 2.9;
2. anterolateral view, x 3.8;
3. dtto, x 3.2;
4. posterolateral view (RA visible), x 3.8;
5. dtto, x 3.1;
Pl. III
1-16 Lecanocrinus soyei Oehlert, 1882
All figures are isolated skeletal elements from the Dvorce-Prokop Limestone at Praha-Hlubočepy, quarry near St. Prokop (Lower Devonian, Pragian, Praha Formation).
1. incomplete IBB circlet with well-observable basal concavity,
L 30574, basal view, x 5;
2. isolated basal, L 30575, outer view, x 5;
3. isolated basal, L 30576, inner view, x 5;
4. isolated SBr, L 30577, outer view, x 5;
5. isolated R, L 30578, outer view, x 3.3;
6. isolated R bearing an undetermined epibiont trace
(pit), L 30579, outer view, x 3.3;
7. extremely thick R, L 30580, inner view, x 3.3;
8. dtto, L 30581, inner view, x 3.3;
9. isolated thick DR, L 30582, inner view, x 4;
10. isolated R, L 30589, outer view, x 4;
11. dtto, L 30584, outer view, x 4;
12. isolated R, L 30585, outer view, x 4;
13. isolated CR with exceptionally distinct surface granulation,
L 30586, outer view, x 2.7;
14. isolated R with partly vermicular sculpture, L 30587,
outer view, x 2.7;
15. isolated CR, L 30588, outer view, 2.7;
16. isolated thick CR, L 30583, inner view, x 2.7;
Pl. IV
1-17 Lecanocrinus soyei Oehlert, 1882
All figures are skeletal elements from the Lower Devonian, Pragian, Praha Formation, "Červený lom" quarry near Praha-Klukovice; 1-8 are derived from the Dvorce-Prokop Limestone, while 9-17 from the Loděnice Limestone.
1. isolated PBr Ax, L 30590, outer view, x 4.3;
2. the same specimen enlarged, x 7.7;
3. isolated SBr, L 30591, distal view, x 4.3;
4. isolated SBr, L 30592, ventral view, x 4.3;
5. isolated BC basal, L 30593, outer view, x 4.3;
6. isolated BC basal, L 30594, inner view, x 4.3;
7. isolated anal X, L 30595, outer view, x 4.3;
8. isolated CR, L 30596, outer view, x 4.3;
9. isolated CD basal, L 30597, outer view, x 3.3;
10. isolated basal, L 30598, outer view, x 3.3;
11. isolated anal X, L 30599, outer view, x 3.3;
12. isolated R, L 30600, outer view, x 3.3;
13. isolated CR, L 30601, outer view, x 3.3;
14. isolated ?CR, L 30602, outer view, x 3.3;
15. isolated R with vermicular sculpture, L 30603, outer
view, x 3.3;
16. the same specimen enlarged, x 6.4;
17. isolated R, L 30626, inner view, x 3.3.
Pl. V
1 Lecanocrinus sp.
Posterolateral view of almost complete crown (L 12106, preserved on weathered surface of a slab of bioclastic limestone with corals), talus zone of the Pragian reef (Lower Devonian, Pragian, Praha Formation, Koněprusy Limestone) at the eastern wall of the "Císařský lom" quarry near Koněprusy. x 5;
2-5 Lecanocrinus hanusi sp. n.
"U kapličky" quarry, Praha-Zlíchov, "Chapel Coral Horizon" at the base of the Zlíchov Limestone (Lower Devonian, Zlíchovian, corresp. to Lower Emsian).
2. aboral cup of the holotype, L 29075, anterolateral
view, x 3.9;
3. dtto, basal view, x 3.1;
4. dtto, posterolateral view (RA missing), x 3.5;
5. CR with PBr1 and PBr Ax attached, L 29076, outer view,
x 4.7;
Pl. VI
1-10 Lecanocrinus hanusi sp. n.
"U kapličky" quarry, Praha-Zlíchov, "Chapel Coral Horizon" at the base of the Zlíchov Limestone (Lower Devonian, Zlíchovian, corresp. to Lower Emsian).
1-3. isolated radial plate with the coiled arm attached
of the paratype A, L 29077, outer to side view, x 3;
4. part of an arm (PBr and SBrBr connected together),
L 29078, inner view showing ventral surface of brachials, x 4.7;
5. a fragment of arms (middle portion) with TBrBr and
QBrBr, L 30564, outer view showing dorsal surface of brachials, x 2.8;
6. one of the typical examples from the rich collections
of F. HanuÜ - two isolated RR and one B glued incorrectly together, L 30604,
x 3.4;
7. isolated DR, L 30605, outer view, x 2;
8. isolated R, L 30606, outer view, x 2;
9. isolated R, L 30607, inner view, x 2;
10. isolated DR, L 30608, inner view, x 2;
Pl. VII
1-10 Lecanocrinus hanusi sp. n.
"U kapličky" quarry, Praha-Zlíchov, "Chapel Coral Horizon" at the base of the Zlíchov Limestone (Lower Devonian, Zlíchovian, corresp. to Lower Emsian).
1. isolated basal, L 30609, inner view, x 2.5;
2. isolated basal, L 30610, outer view, x 2.5;
3. isolated, extremely thick basal, L 30611, inner view,
x 2.4;
4. isolated basal, L 30612, outer view, x 2.4;
5. isolated IB, L 30613, basal view, x 2.5;
6. isolated CR, L 30614, outer view, x 2.2;
7. isolated R, L 30615, outer view, x 2.5;
8. isolated DR, L 30616, outer view, x 2.2;
9. isolated R bearing two specimens of the epibiont trace
Tremichnus sp., L 30617, outer view, x 3.2;
10. isolated CR bearing two specimens of the epibiont
trace Tremichnus sp., L 30618, outer view, x 3.2;
11-17 ??? Lecanocrinus sp.
Praha-Holyně, Třebotov Limestone (Lower Devonian, Dalejan, corresp. to Upper Emsian).
11. isolated ? Anal X, L 30619, outer view, x 6.3;
12. isolated ?CR, L 30620, outer view, x 6.3;
13. isolated R, L 30621, outer view, x 6.3;
14. isolated R, L 30622, inner view, x 7.4;
15. isolated R, L 30623, inner view, x 7.2;
16. isolated ?PBrAx, L 30624, outer view, x 7.4;
17. isolated ?PBrAx, L 30625, distal view, x 7.4.
|
|
|
|
|
|
|